Spinal cord
The spinal cord is a cylindrical structure, greyish-white in colour, connecting the brain to the rest of the body.
The spinal cord arises cranially as a continuation of the medulla oblongata.
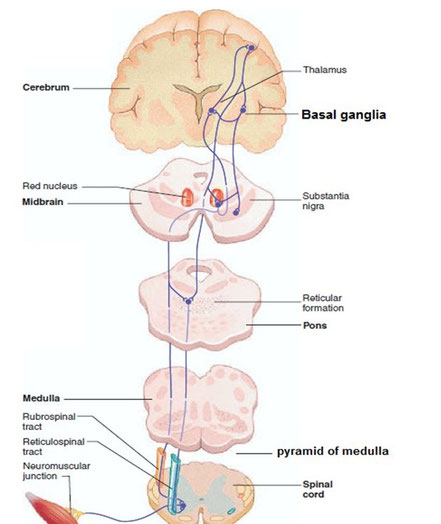
It then travels inferiorly within the vertebral canal, surrounded by the spinal meninges containing cerebrospinal fluid. At roughly the L2 vertebral level the spinal cord tapers off, forming the conus medullaris.
As a result of the termination of the spinal cord at L2, it occupies around two thirds of the vertebral canal. The spinal nerves that arise from the end of the spinal cord are bundled together, forming a structure known as the cauda equina.
During the course of the spinal cord, there are two points of enlargement. The cervical enlargement is located proximally, at the C4-T1 level. It represents the origin of the brachial plexus. Between T11 and L1 is the lumbar enlargement, representing the origin of the lumbar and sacral plexi.
The spinal cord is marked by two depressions on its surface. The anterior median fissure is a deep groove extending the length of the anterior surface of the spinal cord. On the posterior aspect there is a slightly shallower depression – the posterior median sulcus.
Grey matter
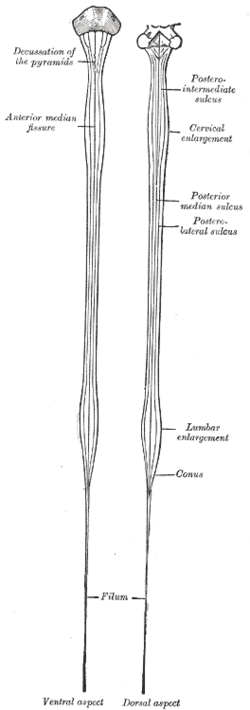
A transverse section of the spinal cord reveals a distinct “butterfly” pattern of dark, inner “grey” matter surrounded by the lighter colour “white matter”.
The grey matter is divided into four main columns: the dorsal horn, the intermediate column, the lateral horn and the ventral horn:
- The dorsal horn (also known as the posterior horn) contains neurons that receive somatosensory information from the body, which is then transmitted via the ascending pathways, to the brain.
- The ventral horn (also known as the anterior horn) largely contains motor neurons that exit the spinal cord to innervate skeletal muscle.
- The intermediate column and lateral horn contains neurons that innervate visceral and pelvic organs.
The prominent nuclei (groups of neuron cell bodies) in the spinal cord are the:
- Marginal zone (MZ, posterior marginalis) – located at the tip of the dorsal horn, and is important for relaying pain and temperature sensation to the brain.
- Substantia gelatinosa (SG) – located at the top of the dorsal horn, the SG is important for relaying pain, temperature and light touch sensation to the brain.
- Nucleus proprius (NP) – located in the ‘neck’ of the dorsal horn, the NP relays mechanical and temperature sensation to the brain.
- Dorsal nucleus of Clarke (DNC) – the most dorso-medial nuclei, the DNC relays unconscious proprioceptive information to the brain. Only found in spinal segments C8 to L3.
- Interomediolateral nucleus (IMN) – located in the intermediate column and lateral horn, the IMN relays sensory information from viscera to the brain, and autonomic signals from the brain to the visceral organs.
- Lateral motor neurons and medial motor neurons (MNs) – located in the ventral horn. Composed of motor neurons that innervate visceral and skeletal muscles.

As an alternative to spinal cord nuclei, Bror Rexed (1950s) identified layers, or laminae, within the spinal cord where cells were grouped according to their structure and function, rather than solely on location.
Lamina I
- tip of the dorsal horn
- cells respond to noxious or thermal stimuli
- sends information to the brain by the contralateral spinothalamic tract
- corresponds to the marginal zone
Lamina II
- Involved in sensation of noxious and non-noxious stimuli, and modulating sensory input to contribute to the brain’s interpretation of incoming signals as painful, or not.
- Sends information to Lamina III and IV
- Corresponds to substantia gelatinosa
Lamina III
- Involved in proprioception and sensation of light touch.
- Cells in this layer connects with cells in layers IV, V and VI.
- Partially corresponds to nucleus proprius
Lamina IV
- Involved in non-noxious sensory information relay and processing.
- Cells connect with those in lamina II
- Partially corresponds to nucleus proprius
Lamina V
- Relays sensory, including nociceptive (potentially painful), information to the brain via the contralateral and spinothalamic tracts
- Receives descending information from the brain via the corticospinal and rubrospinal tracts.
Lamina VI
- Contains many small interneurons involved in spinal reflexes
- Receives sensory information from muscle spindles (involved in proprioception).
- Sends information to the brain via ipsilateral spinocerebellar pathways
Lamina VII
- Large, heterogenous zone that varies through the length of the spinal cord.
- Receives information from Lamina II to VI, and from viscera
- Relays motor information to the viscera
- Gives rise to cells involved in the autonomic system
- Dorsal nucleus of Clarke is part of Lamina VII
Lamina VIII
- Varies depending on spinal cord level, but is most prominent in cervical and lumbar enlargements
- Cells are involved in modulating motor output to skeletal muscle
Lamina IX
- Size and shape varies between spinal cord levels
- Distinct groups of motor neurons that innervate skeletal muscle.
Lamina X
- Surrounds the central canal – the grey commissure
- Axons decussate (cross over) from one side of the spinal cord to the other

White matter
The white matter of the central cord contains the ascending and descending spinal tracts.
The ascending tracts refer to the neural pathways by which sensory information from the peripheral nerves is transmitted to the cerebral cortex. In some texts, ascending tracts are also known as somatosensory pathways or systems.
Ascending tracts
Functionally, the ascending tracts can be divided into the type of information they transmit – conscious or unconscious:
- Conscious tracts – comprised of the dorsal column-medial lemniscal pathway and the anterolateral system.
- Unconscious tracts – comprised of the spinocerebellar tracts.
The dorsal column-medial lemniscal pathway (DCML) carries the sensory modalities of fine touch (tactile sensation), vibration and proprioception.
Its name arises from the two major structures that comprise the DCML. In the spinal cord, information travels via the dorsal (posterior) columns. In the brainstem, it is transmitted through the medial lemniscus.There are three groups of neurones involved in this pathway – first, second and third order neurones.
The first order neurones carry sensory information regarding touch, proprioception or vibration from the peripheral nerves to the medulla oblongata. There are two different pathways which the first order neurones take:
- Signals from the upper limb (T6 and above) – travel in the fasciculus cuneatus (the lateral part of the dorsal column). They then synapse in the nucleus cuneatus of the medulla oblongata.
- Signals from the lower limb (below T6) – travel in the fasciculus gracilis (the medial part of the dorsal column). They then synapse in the nucleus gracilis of the medulla oblongata.
The second order neurones begin in the cuneate nucleus or gracilis. The fibres receive the information from the preceding neurones, and delivers it to the third order neurones in the thalamus. Within the medulla oblongata, these fibres decussate (cross to the other side of the
Lastly, the third order neurones transmit the sensory signals from the thalamus to the ipsilateral primary sensory cortex of the brain. They ascend from the ventral posterolateral nucleus of the thalamus, travel through the internal capsule and terminate at the sensory cortex.
The anterolateral system consists of two separate tracts:
- Anterior spinothalamic tract – carries the sensory modalities of crude touch and pressure.
- Lateral spinothalamic tract – carries the sensory modalities of pain and temperature.
Much like the DCML pathway, both tracts of the anterolateral system have three groups of neurones.
The first order neurones arise from the sensory receptors in the periphery. They enter the spinal cord, ascend 1-2 vertebral levels, and synapse at the tip of the dorsal horn – an area known as the substantia gelatinosa.
The second order neurones carry the sensory information from the substantia gelatinosa to the thalamus. After synapsing with the first order neurones, these fibres decussate within the spinal cord, and then form two distinct tracts:
- Crude touch and pressure fibres – enter the anterior spinothalamic tract.
- Pain and temperature fibres – enter the lateral spinothalamic tract.
Although they are functionally distinct, these tracts run alongside each other, and they can be considered as a single pathway. They travel superiorly within the spinal cord, synapsing in the thalamus.
The third order neurones carry the sensory signals from the thalamus to the ipsilateral primary sensory cortex of the brain. They ascend from the ventral posterolateral nucleus of the thalamus, travel through the internal capsule and terminate at the sensory cortex.
The DCML and the anterolateral tracts transmit conscious sensations, such as pain, touch and temperature. The tracts that carry unconscious proprioceptive information are collectively known as the spinocerebellar tracts.
Although we cannot physically acknowledge these signals, they help our brain co-ordinate and refine motor movements. They transmit information from the muscles to the cerebellum.
Within the spinocerebellar tracts, there are four individual pathways:
- Posterior spinocerebellar tract – Carries proprioceptive information from the lower limbs to the ipsilateral cerebellum.
- Cuneocerebellar tract – Carries proprioceptive information from the upper limbs to the ipsilateral cerebellum.
- Anterior spinocerebellar tract – Carries proprioceptive information from the lower limbs. The fibres decussate twice – and so terminate in the ipsilateral cerebellum.
- Rostral spinocerebellar tract – Carries proprioceptive information from the upper limbs to the ipsilateral cerebellum.
Descending tracts
The descending tracts are the pathways by which motor signals are sent from the brain to lower motor neurones. The lower motor neurones then directly innervate muscles to produce movement.
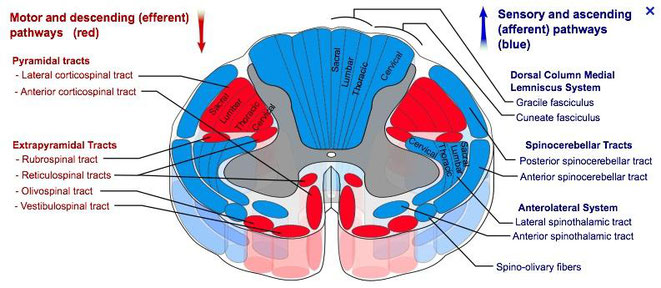
The motor tracts can be functionally divided into two major groups:
- Pyramidal tracts – These tracts originate in the cerebral cortex, carrying motor fibres to the spinal cord and brain stem. They are responsible for the voluntary control of the musculature of the body and face.
- Extrapyramidal tracts – These tracts originate in the brain stem, carrying motor fibres to the spinal cord. They are responsible for the involuntary and automatic control of all musculature, such as muscle tone, balance, posture and locomotion
There are no synapses within the descending pathways. At the termination of the descending tracts, the neurones synapse with a lower motor neurone. Thus, all the neurones within the descending motor system are classed as upper motor neurones. Their cell bodies are found in the cerebral cortex or the brain stem, with their axons remaining within the CNS.
Pyramidal tracts
The pyramidal tracts derive their name from the medullary pyramids of the medulla oblongata, which they pass through. These pathways are responsible for the voluntary control of the musculature of the body and face. Functionally, these tracts can be subdivided into two tracts:
- Corticospinal tracts – supplies the musculature of the body.
- Corticobulbar tracts – supplies the musculature of the head and neck.

The corticospinal tracts begin in the cerebral cortex, from which they receive a range of inputs:
- Primary motor cortex
- Premotor cortex
- Supplementary motor area
They also receive nerve fibres from the somatosensory area, which play a role in regulating the activity of the ascending tracts. After originating from the cortex, the neurones converge, and descend through the internal capsule (a white matter pathway, located between the thalamus and the basal ganglia). This is clinically important, as the internal capsule is particularly susceptible to compression from haemorrhagic bleeds, known as a ‘capsular stroke‘. Such an event could cause a lesion of the descending tracts.
After the internal capsule, the neurones pass through the crus cerebri of the midbrain, the pons and into the medulla. In the most inferior (caudal) part of the medulla, the tract divides into two:
- The fibres within the lateral corticospinal tract decussate (cross over to the other side of the CNS). They then descend into the spinal cord, terminating in the ventral horn (at all segmental levels). From the ventral horn, the lower motor neurones go on to supply the muscles of the body.
- The anterior corticospinal tract remains ipsilateral, descending into the spinal cord. They then decussate and terminate in the ventral horn of the cervical and upper thoracic segmental levels.
The corticobulbar tracts arise from the lateral aspect of the primary motor cortex. They receive the same inputs as the corticospinal tracts. The fibres converge and pass through the internal capsule to the brainstem. The neurones terminate on the motor nuclei of the cranial nerves. Here, they synapse with lower motor neurones, which carry the motor signals to the muscles of the face and neck.
Clinically, it is important to understand the organisation of the corticobulbar fibres. Many of these fibres innervate the motor neurones bilaterally. For example, fibres from the left primary motor cortex act as upper motor neurones for the right and left trochlear nerves. There are a few exceptions to this rule:
- Upper motor neurones for the facial nerve (CN VII) have a contralateral innervation. This only affects the muscles in the lower quadrant of the face – below the
eyes.
- Upper motor neurons for the hypoglossal (CN XII) nerve only provide contralateral innervation.

Extrapyramidal tracts
The extrapyramidal tracts originate in the brainstem, carrying motor fibres to the spinal cord. They are responsible for the involuntary and automatic control of
all musculature, such as muscle tone, balance, posture and locomotion. There are four tracts in total. The vestibulospinal and reticulospinal tracts do not decussate, providing ipsilateral
innervation. The rubrospinal and tectospinal tracts do decussate, and therefore provide contralateral innervation.
Vestibulospinal Tracts
There are two vestibulospinal pathways; medial and lateral. They arise from the vestibular nuclei, which receive input from the organs of balance. The tracts convey this balance information to the spinal cord, where it remains ipsilateral. Fibres in this pathway control balance and posture by innervating the ‘anti-gravity’ muscles (flexors of the arm, and extensors of the leg), via lower motor neurones.
Reticulospinal Tracts
The two recticulospinal tracts have differing functions:
- The medial reticulospinal tract arises from the pons. It facilitates voluntary movements, and increases muscle tone.
- The lateral reticulospinal tract arises from the medulla. It inhibits voluntary movements, and reduces muscle tone.
Rubrospinal Tracts
The rubrospinal tract originates from the red nucleus, a midbrain structure. As the fibres emerge, they decussate (cross over to the other side of the CNS), and
descend into the spinal cord. Thus, they have a contralateral innervation. Its exact function is unclear, but it is thought to play a role in the fine control of hand movements.
Tectospinal Tracts
This pathway begins at the superior colliculus of the midbrain. The superior colliculus is a structure that receives input from the optic nerves. The neurones then quickly decussate, and enter the spinal cord. They terminate at the cervical levels of the spinal cord. The tectospinal tract coordinates movements of the head in relation to vision stimuli.